
We had been studying the role of mountain roads as drivers of non-native plant invasions in mountains all the way back till 2007. Heck, we just published a first manuscript on the decadal dynamics in those (Iseli et al. 2024)! Hiking through the worlds’ beautiful mountains, however, we soon started to wonder: do the same principles hold up along trails as well?
Already back in 2007, we started studying how mountain roads act as highways for non-native plants to invade pristine alpine ecosystems. Heck, we just published a first manuscript on the decadal dynamics in those (Iseli et al. 2025)! Hiking through our beloved mountains all over the world, however, a new question soon emerged:
What about trails?
Trails, like roads, are linear disturbances. They connect valleys with remote mountaintops, they carry humans (and their gear, boots, and seed-laden socks), and they alter local conditions in ways that could favor non-native species – opening up vegetation, increasing nutrient availability, and creating new microhabitats.
But trails are very clearly not roads – I don’t have to tell you that! The disturbance is softer. The infrastructure is minimal – often no gravel, no salt, no foreign materials. Hikers often wander off trail. There’s less edge, less management, and often fewer people. So, we suspected the same mechanisms would play out, but at a lower intensity. Fewer non-natives, lower cover, and smaller contrasts between trail edges and adjacent vegetation.
And so, an idea was born: take the MIREN road survey protocol and adapt it for trails.

In 2016, without any dedicated funding – just shared enthusiasm and a love for mountains – we began surveying trails worldwide. Today, thanks to a remarkable effort relentlessly led by Agustina Barros and Eduardo Fuentes Lillo, we’ve surveyed 55 trails across 9 regions, and the first results are now published in Global Ecology and Biogeography.
So, what did we find?
At the global scale, climate (especially temperature) explained most of the variation in non-native plant richness. Biotic factors – like the diversity of native vegetation – best predicted presence and cover. Human-related factors (trail use intensity, grazing, distance to the trail) also played a role, but less consistently.
Importantly, the patterns were strikingly similar to those we’ve seen along roads:
- Fewer non-natives at higher elevations and latitudes
- Fewer non-natives farther from the trail edge
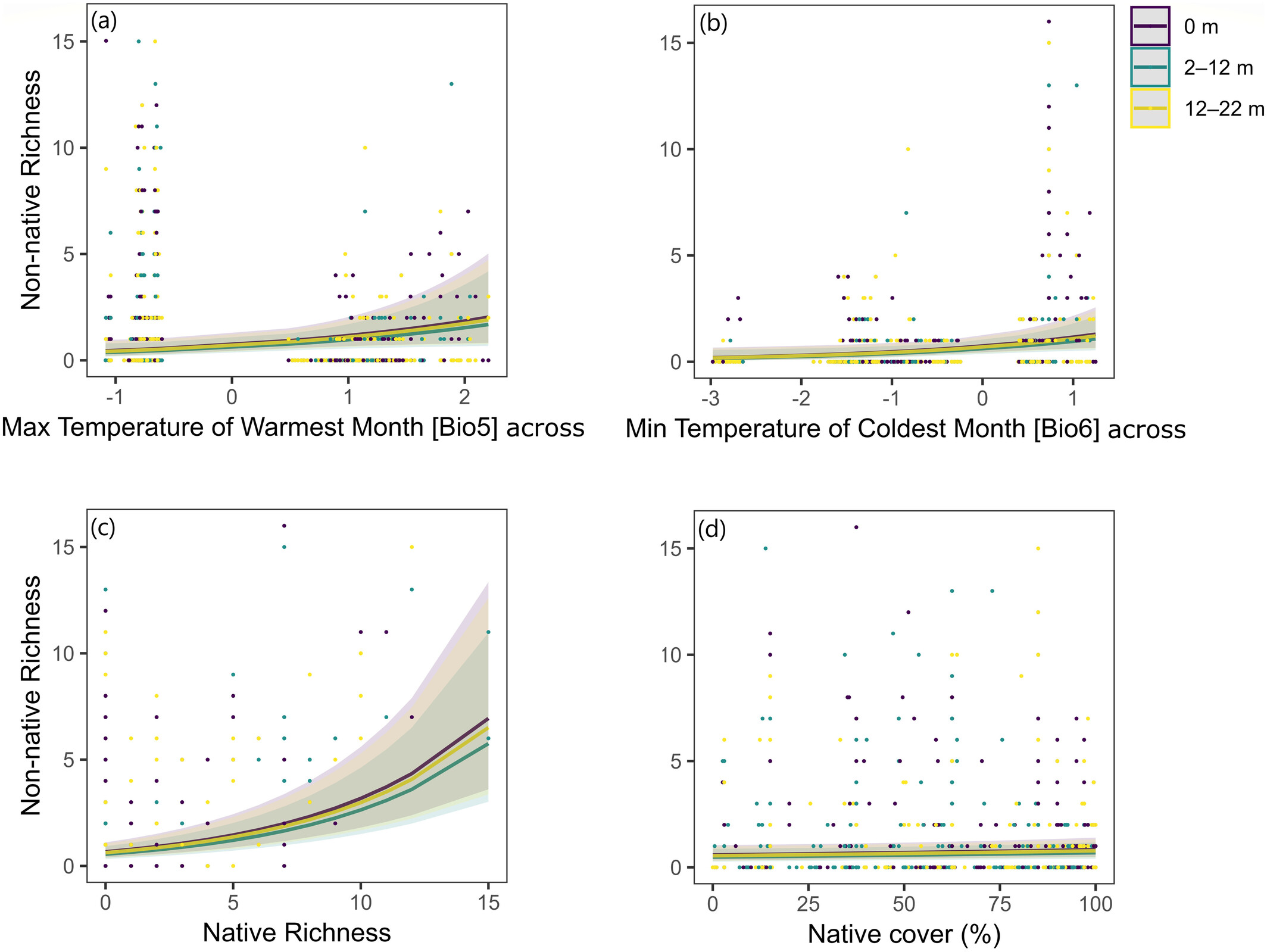
This supports the idea of a “double filter” for non-native plants:
- Climate limits which species can survive as you go higher and colder.
- Distance from disturbance limits where they can establish, as native communities become more resilient, and propagule availability lower.
As expected, however, the impact of trails was indeed less intense than that of roads!

What’s also fascinating is that the same few non-native species – often hardy European forbs like Taraxacum officinale, Trifolium repens, and Rumex acetosella – pop up again and again, regardless of continent. These generalist species are good at exploiting disturbed habitats and are likely part of the global alpine invasion front.

So, should we worry?
Most of the non-native species we observed are not yet invasive. And we certainly recognize the many benefits of trails – for people, for recreation, and even for biodiversity in some contexts.

But education matters. Encouraging hikers to stay on marked trails, limiting the expansion of trail networks, and raising awareness about their role in shaping ecosystems can help reduce further spread. Most of all, we need to focus on prevention – especially since the coldest, highest-elevation areas still remain relatively free of non-native species.
And while we may not stop every seed from catching a ride up the trail, this work shows we can still limit where it lands, and whether it takes root.
Reference: Barros et al. (2025). Beyond the Trail—Understanding Non-Native Plant Invasions in Mountain Ecosystems. Global Ecology and Biogeography. http://dx.doi.org/10.1111/geb.70060











































































































































































































































































































































































































































































































































































































































































































































































































































































")
















")
")





")




































































































")




































































")



















































")













")
































































